Messenger RNA

 Clash Royale CLAN TAG#URR8PPP
Clash Royale CLAN TAG#URR8PPP 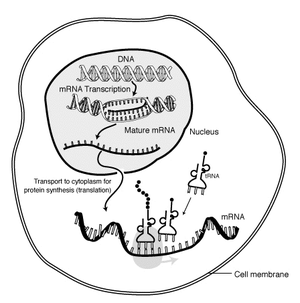
The "life cycle" of an mRNA in a eukaryotic cell. RNA is transcribed in the nucleus; after processing, it is transported to the cytoplasm and translated by the ribosome. Finally, the mRNA is degraded.
Messenger RNA (mRNA) is a large family of RNA molecules that convey genetic information from DNA to the ribosome, where they specify the amino acid sequence of the protein products of gene expression. RNA polymerase transcribes primary transcript mRNA (known as pre-mRNA) into processed, mature mRNA. This mature mRNA is then translated into a polymer of amino acids: a protein, as summarized in the central dogma of molecular biology.
As in DNA, mRNA genetic information is in the sequence of nucleotides, which are arranged into codons consisting of three base pairs each. Each codon encodes for a specific amino acid, except the stop codons, which terminate protein synthesis. This process of translation of codons into amino acids requires two other types of RNA: Transfer RNA (tRNA), that mediates recognition of the codon and provides the corresponding amino acid, and ribosomal RNA (rRNA), that is the central component of the ribosome's protein-manufacturing machinery.
The existence of mRNA was first suggested by Jacques Monod and François Jacob, and subsequently discovered by Jacob, Sydney Brenner and Matthew Meselson at the California Institute of Technology in 1961.
It should not be confused with mitochondrial DNA.
Contents
1 Synthesis, processing and function
1.1 Transcription
1.2 Eukaryotic pre-mRNA processing
1.2.1 5' cap addition
1.2.2 Editing
1.2.3 Polyadenylation
1.3 Transport
1.4 Translation
2 Structure
2.1 Coding regions
2.2 Untranslated regions
2.3 Poly(A) tail
2.4 Monocistronic versus polycistronic mRNA
2.5 mRNA circularization
3 Degradation
3.1 Prokaryotic mRNA degradation
3.2 Eukaryotic mRNA turnover
3.3 AU-rich element decay
3.4 Nonsense mediated decay
3.5 Small interfering RNA (siRNA)
3.6 MicroRNA (miRNA)
3.7 Other decay mechanisms
4 mRNA-based therapeutics
5 Further reading
6 See also
7 References
8 External links
Synthesis, processing and function
The brief existence of an mRNA molecule begins with transcription, and ultimately ends in degradation. During its life, an mRNA molecule may also be processed, edited, and transported prior to translation. Eukaryotic mRNA molecules often require extensive processing and transport, while prokaryotic mRNA molecules do not. A molecule of eukaryotic mRNA and the proteins surrounding it are together called a messenger RNP.
Transcription
Transcription is when RNA is made from DNA. During transcription, RNA polymerase makes a copy of a gene from the DNA to mRNA as needed. This process is similar in eukaryotes and prokaryotes. One notable difference, however, is that eukaryotic RNA polymerase associates with mRNA-processing enzymes during transcription so that processing can proceed quickly after the start of transcription. The short-lived, unprocessed or partially processed product is termed precursor mRNA, or pre-mRNA; once completely processed, it is termed mature mRNA.
Eukaryotic pre-mRNA processing
Processing of mRNA differs greatly among eukaryotes, bacteria, and archea. Non-eukaryotic mRNA is, in essence, mature upon transcription and requires no processing, except in rare cases. Eukaryotic pre-mRNA, however, requires extensive processing.
5' cap addition
A 5' cap (also termed an RNA cap, an RNA 7-methylguanosine cap, or an RNA m7G cap) is a modified guanine nucleotide that has been added to the "front" or 5' end of a eukaryotic messenger RNA shortly after the start of transcription. The 5' cap consists of a terminal 7-methylguanosine residue that is linked through a 5'-5'-triphosphate bond to the first transcribed nucleotide. Its presence is critical for recognition by the ribosome and protection from RNases.
Cap addition is coupled to transcription, and occurs co-transcriptionally, such that each influences the other. Shortly after the start of transcription, the 5' end of the mRNA being synthesized is bound by a cap-synthesizing complex associated with RNA polymerase. This enzymatic complex catalyzes the chemical reactions that are required for mRNA capping. Synthesis proceeds as a multi-step biochemical reaction.
Editing
In some instances, an mRNA will be edited, changing the nucleotide composition of that mRNA. An example in humans is the apolipoprotein B mRNA, which is edited in some tissues, but not others. The editing creates an early stop codon, which, upon translation, produces a shorter protein.
Polyadenylation
Polyadenylation is the covalent linkage of a polyadenylyl moiety to a messenger RNA molecule. In eukaryotic organisms most messenger RNA (mRNA) molecules are polyadenylated at the 3' end, but recent studies have shown that short stretches of uridine (oligouridylation) are also common.[1] The poly(A) tail and the protein bound to it aid in protecting mRNA from degradation by exonucleases. Polyadenylation is also important for transcription termination, export of the mRNA from the nucleus, and translation. mRNA can also be polyadenylated in prokaryotic organisms, where poly(A) tails act to facilitate, rather than impede, exonucleolytic degradation.
Polyadenylation occurs during and/or immediately after transcription of DNA into RNA. After transcription has been terminated, the mRNA chain is cleaved through the action of an endonuclease complex associated with RNA polymerase. After the mRNA has been cleaved, around 250 adenosine residues are added to the free 3' end at the cleavage site. This reaction is catalyzed by polyadenylate polymerase. Just as in alternative splicing, there can be more than one polyadenylation variant of an mRNA.
Polyadenylation site mutations also occur. The primary RNA transcript of a gene is cleaved at the poly-A addition site, and 100-200 A's are added to the 3’ end of the RNA. If this site is altered, an abnormally long and unstable mRNA construct will be formed.
Transport
Another difference between eukaryotes and prokaryotes is mRNA transport. Because eukaryotic transcription and translation is compartmentally separated, eukaryotic mRNAs must be exported from the nucleus to the cytoplasm—a process that may be regulated by different signaling pathways.[2] Mature mRNAs are recognized by their processed modifications and then exported through the nuclear pore by binding to the cap-binding proteins CBP20 and CBP80,[3] as well as the transcription/export complex (TREX).[4][5] Multiple mRNA export pathways have been identified in eukaryotes.[6]
In spatially complex cells, some mRNAs are transported to particular subcellar destinations. In mature neurons, certain mRNA are transported from the soma to dendrites. One site of mRNA translation is at polyribosomes selectively localized beneath synapses.[7] The mRNA for Arc/Arg3.1 is induced by synaptic activity and localizes selectively near active synapses based on signals generated by NMDA receptors.[8] Other mRNAs also move into dendrites in response to external stimuli, such as β-actin mRNA.[9] Upon export from the nucleus, actin mRNA associates with ZBP1 and the 40S subunit. The complex is bound by a motor protein and is transported to the target location (neurite extension) along the cytoskeleton. Eventually ZBP1 is phosphorylated by Src in order for translation to be initiated.[10] In developing neurons, mRNAs are also transported into growing axons and especially growth cones. Many mRNAs are marked with so-called "zip codes," which target their transport to a specific location.[11]
Translation
Because prokaryotic mRNA does not need to be processed or transported, translation by the ribosome can begin immediately after the end of transcription. Therefore, it can be said that prokaryotic translation is coupled to transcription and occurs co-transcriptionally.
Eukaryotic mRNA that has been processed and transported to the cytoplasm (i.e., mature mRNA) can then be translated by the ribosome. Translation may occur at ribosomes free-floating in the cytoplasm, or directed to the endoplasmic reticulum by the signal recognition particle. Therefore, unlike in prokaryotes, eukaryotic translation is not directly coupled to transcription.[12] It is even possible in some contexts that reduced mRNA levels are accompanied by increased protein levels, as has been observed for mRNA/protein levels of EEF1A1 in breast cancer.[13]
Structure

The structure of a mature eukaryotic mRNA. A fully processed mRNA includes a 5' cap, 5' UTR, coding region, 3' UTR, and poly(A) tail.
Coding regions
Coding regions are composed of codons, which are decoded and translated (in eukaryotes usually into one and in prokaryotes usually into several) into proteins by the ribosome. Coding regions begin with the start codon and end with a stop codon. In general, the start codon is an AUG triplet and the stop codon is UAA, UAG, or UGA. The coding regions tend to be stabilised by internal base pairs, this impedes degradation.[14][15] In addition to being protein-coding, portions of coding regions may serve as regulatory sequences in the pre-mRNA as exonic splicing enhancers or exonic splicing silencers.
Untranslated regions
Untranslated regions (UTRs) are sections of the mRNA before the start codon and after the stop codon that are not translated, termed the five prime untranslated region (5' UTR) and three prime untranslated region (3' UTR), respectively. These regions are transcribed with the coding region and thus are exonic as they are present in the mature mRNA. Several roles in gene expression have been attributed to the untranslated regions, including mRNA stability, mRNA localization, and translational efficiency. The ability of a UTR to perform these functions depends on the sequence of the UTR and can differ between mRNAs. Genetic variants in 3' UTR have also been implicated in disease susceptibility because of the change in RNA structure and protein translation.[16]
The stability of mRNAs may be controlled by the 5' UTR and/or 3' UTR due to varying affinity for RNA degrading enzymes called ribonucleases and for ancillary proteins that can promote or inhibit RNA degradation. (See also, C-rich stability element.)
Translational efficiency, including sometimes the complete inhibition of translation, can be controlled by UTRs. Proteins that bind to either the 3' or 5' UTR may affect translation by influencing the ribosome's ability to bind to the mRNA. MicroRNAs bound to the 3' UTR also may affect translational efficiency or mRNA stability.
Cytoplasmic localization of mRNA is thought to be a function of the 3' UTR. Proteins that are needed in a particular region of the cell can also be translated there; in such a case, the 3' UTR may contain sequences that allow the transcript to be localized to this region for translation.
Some of the elements contained in untranslated regions form a characteristic secondary structure when transcribed into RNA. These structural mRNA elements are involved in regulating the mRNA. Some, such as the SECIS element, are targets for proteins to bind. One class of mRNA element, the riboswitches, directly bind small molecules, changing their fold to modify levels of transcription or translation. In these cases, the mRNA regulates itself.
Poly(A) tail
The 3' poly(A) tail is a long sequence of adenine nucleotides (often several hundred) added to the 3' end of the pre-mRNA. This tail promotes export from the nucleus and translation, and protects the mRNA from degradation.
Monocistronic versus polycistronic mRNA
An mRNA molecule is said to be monocistronic when it contains the genetic information to translate only a single protein chain (polypeptide). This is the case for most of the eukaryotic mRNAs.[17][18] On the other hand, polycistronic mRNA carries several open reading frames (ORFs), each of which is translated into a polypeptide. These polypeptides usually have a related function (they often are the subunits composing a final complex protein) and their coding sequence is grouped and regulated together in a regulatory region, containing a promoter and an operator. Most of the mRNA found in bacteria and archaea is polycistronic,[17] as is the human mitochondrial genome.[19] Dicistronic or bicistronic mRNA encodes only two proteins.
mRNA circularization
In eukaryotes mRNA molecules form circular structures due to an interaction between the eIF4E and poly(A)-binding protein, which both bind to eIF4G, forming an mRNA-protein-mRNA bridge.[20] Circularization is thought to promote cycling of ribosomes on the mRNA leading to time-efficient translation, and may also function to ensure only intact mRNA are translated (partially degraded mRNA characteristically have no m7G cap, or no poly-A tail).[21]
Other mechanisms for circularization exist, particularly in virus mRNA. Poliovirus mRNA uses a cloverleaf section towards its 5' end to bind PCBP2, which binds poly(A)-binding protein, forming the familiar mRNA-protein-mRNA circle. Barley yellow dwarf virus has binding between mRNA segments on its 5' end and 3' end (called kissing stem loops), circularizing the mRNA without any proteins involved.
RNA virus genomes (the + strands of which are translated as mRNA) are also commonly circularized.[citation needed] During genome replication the circularization acts to enhance genome replication speeds, cycling viral RNA-dependent RNA polymerase much the same as the ribosome is hypothesized to cycle.
Degradation
Different mRNAs within the same cell have distinct lifetimes (stabilities). In bacterial cells, individual mRNAs can survive from seconds to more than an hour. However, the lifetime averages between 1 and 3 minutes, making bacterial mRNA much less stable than eukaryotic mRNA.[22] In mammalian cells, mRNA lifetimes range from several minutes to days.[23] The greater the stability of an mRNA the more protein may be produced from that mRNA. The limited lifetime of mRNA enables a cell to alter protein synthesis rapidly in response to its changing needs. There are many mechanisms that lead to the destruction of an mRNA, some of which are described below.
Prokaryotic mRNA degradation
In general, in prokaryotes the lifetime of mRNA is much shorter than in eukaryotes. Prokaryotes degrade messages by using a combination of ribonucleases, including endonucleases, 3' exonucleases, and 5' exonucleases. In some instances, small RNA molecules (sRNA) tens to hundreds of nucleotides long can stimulate the degradation of specific mRNAs by base-pairing with complementary sequences and facilitating ribonuclease cleavage by RNase III. It was recently shown that bacteria also have a sort of 5' cap consisting of a triphosphate on the 5' end.[24] Removal of two of the phosphates leaves a 5' monophosphate, causing the message to be destroyed by the exonuclease RNase J, which degrades 5' to 3'.
Eukaryotic mRNA turnover
Inside eukaryotic cells, there is a balance between the processes of translation and mRNA decay. Messages that are being actively translated are bound by ribosomes, the eukaryotic initiation factors eIF-4E and eIF-4G, and poly(A)-binding protein. eIF-4E and eIF-4G block the decapping enzyme (DCP2), and poly(A)-binding protein blocks the exosome complex, protecting the ends of the message. The balance between translation and decay is reflected in the size and abundance of cytoplasmic structures known as P-bodies[25] The poly(A) tail of the mRNA is shortened by specialized exonucleases that are targeted to specific messenger RNAs by a combination of cis-regulatory sequences on the RNA and trans-acting RNA-binding proteins. Poly(A) tail removal is thought to disrupt the circular structure of the message and destabilize the cap binding complex. The message is then subject to degradation by either the exosome complex or the decapping complex. In this way, translationally inactive messages can be destroyed quickly, while active messages remain intact. The mechanism by which translation stops and the message is handed-off to decay complexes is not understood in detail.
AU-rich element decay
The presence of AU-rich elements in some mammalian mRNAs tends to destabilize those transcripts through the action of cellular proteins that bind these sequences and stimulate poly(A) tail removal. Loss of the poly(A) tail is thought to promote mRNA degradation by facilitating attack by both the exosome complex[26] and the decapping complex.[27] Rapid mRNA degradation via AU-rich elements is a critical mechanism for preventing the overproduction of potent cytokines such as tumor necrosis factor (TNF) and granulocyte-macrophage colony stimulating factor (GM-CSF).[28] AU-rich elements also regulate the biosynthesis of proto-oncogenic transcription factors like c-Jun and c-Fos.[29]
Nonsense mediated decay
Eukaryotic messages are subject to surveillance by nonsense mediated decay (NMD), which checks for the presence of premature stop codons (nonsense codons) in the message. These can arise via incomplete splicing, V(D)J recombination in the adaptive immune system, mutations in DNA, transcription errors, leaky scanning by the ribosome causing a frame shift, and other causes. Detection of a premature stop codon triggers mRNA degradation by 5' decapping, 3' poly(A) tail removal, or endonucleolytic cleavage.[30]
Small interfering RNA (siRNA)
In metazoans, small interfering RNAs (siRNAs) processed by Dicer are incorporated into a complex known as the RNA-induced silencing complex or RISC. This complex contains an endonuclease that cleaves perfectly complementary messages to which the siRNA binds. The resulting mRNA fragments are then destroyed by exonucleases. siRNA is commonly used in laboratories to block the function of genes in cell culture. It is thought to be part of the innate immune system as a defense against double-stranded RNA viruses.[31]
MicroRNA (miRNA)
MicroRNAs (miRNAs) are small RNAs that typically are partially complementary to sequences in metazoan messenger RNAs.[32] Binding of a miRNA to a message can repress translation of that message and accelerate poly(A) tail removal, thereby hastening mRNA degradation. The mechanism of action of miRNAs is the subject of active research.[33]
Other decay mechanisms
There are other ways by which messages can be degraded, including non-stop decay and silencing by Piwi-interacting RNA (piRNA), among others.
mRNA-based therapeutics
Full length mRNA molecules have been proposed as therapeutics since the beginning of the biotech era but there was little traction until the 2010s, when Moderna Therapeutics was founded and managed to raise almost a billion dollars in venture funding in its first three years.[34][35][36][37]
Theoretically, the administered mRNA sequence can cause a cell to make a protein, which in turn could directly treat a disease or could function as a vaccine; more indirectly the protein could drive an endogenous stem cell to differentiate in a desired way.[38][39]
The primary challenges of RNA therapy center on delivering the RNA to directed cells, more even than determining what sequence to deliver.[35] Naked RNA sequences will naturally degrade after preparation; they may trigger the body's immune system to attack them as an invader; and they are impermeable to the cell membrane.[39] Once within the cell, they must then leave the cell's transport mechanism to take action within the cytoplasm, which houses the ribosomes that direct manufacture of proteins.[38]
Further reading
Alsaweed, Mohammed; Lai, Ching Tat; Hartmann, Peter E.; Geddes, Donna T.; Kakulas, Foteini (8 February 2016). "Human milk miRNAs primarily originate from the mammary gland resulting in unique miRNA profiles of fractionated milk". Scientific Reports. 6 (1). doi:10.1038/srep20680..mw-parser-output cite.citationfont-style:inherit.mw-parser-output qquotes:"""""""'""'".mw-parser-output code.cs1-codecolor:inherit;background:inherit;border:inherit;padding:inherit.mw-parser-output .cs1-lock-free abackground:url("//upload.wikimedia.org/wikipedia/commons/thumb/6/65/Lock-green.svg/9px-Lock-green.svg.png")no-repeat;background-position:right .1em center.mw-parser-output .cs1-lock-limited a,.mw-parser-output .cs1-lock-registration abackground:url("//upload.wikimedia.org/wikipedia/commons/thumb/d/d6/Lock-gray-alt-2.svg/9px-Lock-gray-alt-2.svg.png")no-repeat;background-position:right .1em center.mw-parser-output .cs1-lock-subscription abackground:url("//upload.wikimedia.org/wikipedia/commons/thumb/a/aa/Lock-red-alt-2.svg/9px-Lock-red-alt-2.svg.png")no-repeat;background-position:right .1em center.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registrationcolor:#555.mw-parser-output .cs1-subscription span,.mw-parser-output .cs1-registration spanborder-bottom:1px dotted;cursor:help.mw-parser-output .cs1-hidden-errordisplay:none;font-size:100%.mw-parser-output .cs1-visible-errorfont-size:100%.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration,.mw-parser-output .cs1-formatfont-size:95%.mw-parser-output .cs1-kern-left,.mw-parser-output .cs1-kern-wl-leftpadding-left:0.2em.mw-parser-output .cs1-kern-right,.mw-parser-output .cs1-kern-wl-rightpadding-right:0.2em
Lillycrop, Karen A.; Burdge, Graham C. (October 2012). "Epigenetic mechanisms linking early nutrition to long term health". Best Practice & Research Clinical Endocrinology & Metabolism. 26 (5): 667–676. doi:10.1016/j.beem.2012.03.009. PMID 22980048.
Melnik, Bodo C.; Kakulas, Foteini; Geddes, Donna T.; Hartmann, Peter E.; John, Swen Malte; Carrera-Bastos, Pedro; Cordain, Loren; Schmitz, Gerd (21 June 2016). "Milk miRNAs: simple nutrients or systemic functional regulators?". Nutrition & Metabolism. 13 (1): 42. doi:10.1186/s12986-016-0101-2. PMC 4915038. PMID 27330539.
Vickers, Mark (2 June 2014). "Early Life Nutrition, Epigenetics and Programming of Later Life Disease". Nutrients. 6 (6): 2165–2178. doi:10.3390/nu6062165. PMC 4073141. PMID 24892374.
Zhou, Q; Li, M; Wang, X; Li, Q; Wang, T; Zhu, Q; Zhou, X; Wang, X; Gao, X; Li, X (2012). "Immune-related microRNAs are abundant in breast milk exosomes". International Journal of Biological Sciences. 8 (1): 118–23. PMC 3248653. PMID 22211110.
See also
GeneCalling, an mRNA profiling technology
Transcriptome, the sum of all RNA in a cell
References
^ Choi et al. RNA. 2012. 18: 394-401
^ Quaresma, Alexandre J.; Sievert, Jeffrey A.; Nickerson, J. A. (2013), "Regulation of mRNA export by the PI3 kinase/AKT signal transduction pathway", Mol Biol Cell, 24 (8): 1208–21, doi:10.1091/mbc.E12-06-0450, PMC 3623641, PMID 23427269
^ Kierzkowski, Daniel; Kmieciak, Maciej; Piontek, Paulina; Wojtaszek, Przemyslaw; Szweykowska-Kulinska, Zofia; Jarmolowski, Artur (September 2009). "The Arabidopsis CBP20 targets the cap-binding complex to the nucleus, and is stabilized by CBP80". The Plant Journal. 59 (5): 814–825. doi:10.1111/j.1365-313X.2009.03915.x. Retrieved 12 December 2014.
^ Sträßer, Katja; Masuda, Seiji; Mason, Paul; Pfannstiel, Jens; Oppizzi, Marisa; Rodriguez-Navarro, Susana; Rondón, Ana G.; Aguilera, Andres; Struhl, Kevin; Reed, Robin; Hurt, Ed (28 April 2002). "TREX is a conserved complex coupling transcription with messenger RNA export". Nature. 417 (6886): 304–308. doi:10.1038/nature746. PMID 11979277. Retrieved 12 December 2014.
^ Katahira, Jun; Yoneda, Yoshihiro (27 October 2014). "Roles of the TREX complex in nuclear export of mRNA". RNA Biology. 6 (2): 149–152. doi:10.4161/rna.6.2.8046. Retrieved 12 December 2014.
^ Cenik, Can; Chua, Hon Nian; Zhang, Hui; Tarnawsky, Stefan P.; Akef, Abdalla; Derti, Adnan; Tasan, Murat; Moore, Melissa J.; Palazzo, Alexander F.; Roth, Frederick P. (2011). "Genome Analysis Reveals Interplay between 5′UTR Introns and Nuclear mRNA Export for Secretory and Mitochondrial Genes". PLoS Genetics. 7 (4): e1001366. doi:10.1371/journal.pgen.1001366. ISSN 1553-7404. PMC 3077370. PMID 21533221.
^ Steward, Oswald. "Preferential localization of polyribosomes under the base of dendritic spines in granule cells of the dentate gyrus". Journal of Neuroscience. 2: 284–291.
^ Steward, Oswald (2001). "Selective targeting of newly-synthesized Arc mRNA to active synapses requires NMDA receptor activation". Neuron. 30 (1): 227–240. doi:10.1016/s0896-6273(01)00275-6. PMID 11343657.
^ Job, C.; Eberwine, J. (1912), "Localization and translation of mRNA in dendrites and axons", Nat Rev Neurosci, 2 (12): 889–98, doi:10.1038/35104069, PMID 11733796
^ Spatial regulation of bold beta-actin translation by Src-dependent phosphorylation of ZBP1 Nature04115.
^ Ainger, Kevin; Avossa, Daniela; Diana, Amy S.; Barry, Christopher; Barbarese, Elisa; Carson, John H. (1997), "Transport and Localization Elements in Myelin Basic Protein mRNA", The Journal of Cell Biology, 138 (5): 1077–1087, doi:10.1083/jcb.138.5.1077, PMC 2136761, PMID 9281585
^ somalia
^ Lin, C.Y.; Beattie, A.; Baradaran, B.; Dray, E.; Duijf, P.H.G. (2018), "Contradictory mRNA and protein misexpression of EEF1A1 in ductal breast carcinoma due to cell cycle regulation and cellular stress", Scientific Reports, 8 (1): 13904, doi:10.1038/s41598-018-32272-x, PMID 30224719
^ Shabalina SA, Ogurtsov AY, Spiridonov NA (2006), "A periodic pattern of mRNA secondary structure created by the genetic code", Nucleic Acids Res., 34 (8): 2428–37, doi:10.1093/nar/gkl287, PMC 1458515, PMID 16682450
^ Katz L, Burge CB (September 2003), "Widespread Selection for Local RNA Secondary Structure in Coding Regions of Bacterial Genes", Genome Res., 13 (9): 2042–51, doi:10.1101/gr.1257503, PMC 403678, PMID 12952875
^ Lu, YF; Mauger, DM; Goldstein, DB; Urban, TJ; Weeks, KM; Bradrick, SS (4 November 2015). "IFNL3 mRNA structure is remodeled by a functional non-coding polymorphism associated with hepatitis C virus clearance". Scientific Reports. 5: 16037. doi:10.1038/srep16037. PMC 4631997. PMID 26531896.
^ ab
Kozak, M. (March 1983), "Comparison of initiation of protein synthesis in procaryotes, eucaryotes, and organelles", Microbiological Reviews, 47 (1): 1–45, PMC 281560, PMID 6343825
^ Niehrs C, Pollet N (December 1999), "Synexpression groups in eukaryotes", Nature, 402 (6761): 483–7, doi:10.1038/990025, PMID 10591207
^ Mercer, Tim R.; Neph, Shane; Dinger, Marcel E.; Crawford, Joanna; Smith, Martin A.; Shearwood, Anne-Marie J.; Haugen, Eric; Bracken, Cameron P.; Rackham, Oliver; Stamatoyannopoulos, John A.; Filipovska, Aleksandra; Mattick, John S. (2011). "The Human Mitochondrial Transcriptome". Cell. 146 (4): 645–658. doi:10.1016/j.cell.2011.06.051. ISSN 0092-8674. PMC 3160626. PMID 21854988.
^ Wells, S.E.; Hillner, P.E.; Vale, R.D.; Sachs, A.B. (1998), "Circularization of mRNA by Eukaryotic Translation Initiation Factors" (w), Molecular Cell, 2 (1): 135–140, doi:10.1016/S1097-2765(00)80122-7, PMID 9702200
^ López-Lastra M, Rivas A, Barría MI (2005), "Protein synthesis in eukaryotes: The growing biological relevance of cap-independent translation initiation", Biological Research, 38 (2–3): 121–146, doi:10.4067/S0716-97602005000200003, PMID 16238092CS1 maint: Uses authors parameter (link)
^ Lewin's genes X. Lewin, Benjamin., Krebs, Jocelyn E., Kilpatrick, Stephen T., Goldstein, Elliott S., Lewin, Benjamin. (10th ed.). Sudbury, Mass.: Jones and Bartlett. 2011. ISBN 9780763766320. OCLC 456641931.
^ Yu, Jia; Russell, J. Eric (2001). "Structural and Functional Analysis of an mRNP Complex That Mediates the High Stability of Human β-Globin mRNA" (PDF). Mol Cell Biol. 21 (17): 5879–88. doi:10.1128/mcb.21.17.5879-5888.2001. PMC 87307. PMID 11486027. Retrieved 4 June 2014.
^ Deana, Atilio; Celesnik, Helena; Belasco, Joel G. (2008), "The bacterial enzyme RppH triggers messenger RNA degradation by 5' pyrophosphate removal", Nature, 451 (7176): 355–8, doi:10.1038/nature06475, PMID 18202662
^ Parker, R.; Sheth, U. (2007), "P Bodies and the Control of mRNA Translation and Degradation" (w), Molecular Cell, 25 (5): 635–646, doi:10.1016/j.molcel.2007.02.011, PMID 17349952
^ Chen, C.Y.; Gherzi, R.; Ong, S.E.; Chan, E.L.; Raijmakers, R.; Pruijn, G.J.M.; Stoecklin, G.; Moroni, C.; Mann, M.; Karin, Michael (2001), "AU Binding Proteins Recruit the Exosome to Degrade ARE-Containing mRNAs", Cell, 107 (4): 451–464, doi:10.1016/S0092-8674(01)00578-5, PMID 11719186
^ Fenger-Grøn M, Fillman C, Norrild B, Lykke-Andersen J (December 2005), "Multiple processing body factors and the ARE binding protein TTP activate mRNA decapping" (PDF), Mol. Cell, 20 (6): 905–15, doi:10.1016/j.molcel.2005.10.031, PMID 16364915, archived from the original (PDF) on 2011-06-06
^ Shaw G, Kamen R (August 1986), "A conserved AU sequence from the 3' untranslated region of GM-CSF mRNA mediates selective mRNA degradation", Cell, 46 (5): 659–67, doi:10.1016/0092-8674(86)90341-7, PMID 3488815
^ Chen, C.Y.A.; Shyu, A.B. (1995), "AU-rich elements: characterization and importance in mRNA degradation", Trends in Biochemical Sciences, 20 (11): 465–470, doi:10.1016/S0968-0004(00)89102-1, PMID 8578590
^ Isken, O.; Maquat, L.E. (2007), "Quality control of eukaryotic mRNA: safeguarding cells from abnormal mRNA function", Genes & Development, 21 (15): 1833–56, doi:10.1101/gad.1566807, PMID 17671086
^ Obbard, D.J.; Gordon, K.H.J.; Buck, A.H.; Jiggins, F.M. (2009), "The evolution of RNAi as a defence against viruses and transposable elements", Philosophical Transactions of the Royal Society B: Biological Sciences, 364 (1513): 99–115, doi:10.1098/rstb.2008.0168, PMC 2592633, PMID 18926973
^ Brennecke J, Stark A, Russell RB, Cohen SM (March 2005), "Principles of MicroRNA–Target Recognition", PLoS Biol., 3 (3): e85, doi:10.1371/journal.pbio.0030085, PMC 1043860, PMID 15723116
^ Eulalio, A.; Huntzinger, E.; Nishihara, T.; Rehwinkel, J.; Fauser, M.; Izaurralde, E. (2009), "Deadenylation is a widespread effect of miRNA regulation", RNA, 15 (1): 21–32, doi:10.1261/rna.1399509, PMC 2612776, PMID 19029310
^ Gregory Huang (December 6, 2012). "Moderna, $40M in Tow, Hopes to Reinvent Biotech with "Make Your Own Drug"". Xconomy.
^ ab Kaczmarek, JC; Kowalski, PS; Anderson, DG (27 June 2017). "Advances in the delivery of RNA therapeutics: from concept to clinical reality". Genome medicine. 9 (1): 60. doi:10.1186/s13073-017-0450-0. PMC 5485616. PMID 28655327.
^ Crow, David (7 September 2017). "Moderna pledge sets investor pulses racing; Biotech group promises to rewrite rules of drug development - though sceptics say $5bn valuation is undeserved". Financial Times.
^ Adams, Ben (September 14, 2017). "Moderna changes up R&D model, posts cancer, CV trial updates". FierceBiotech.
^ ab Hajj, Khalid A.; Whitehead, Kathryn A. (12 September 2017). "Tools for translation: non-viral materials for therapeutic mRNA delivery". Nature Reviews Materials. 2 (10): 17056. doi:10.1038/natrevmats.2017.56.
^ ab Gousseinov, Elina; Kozlov, Mikhail; Scanlan, Claire; Hammons, Aaron; Bei, Ling; Benchek, Youssef; Pirani, Karim; Pattnaik, Priyabrata; Waghmare, Ruta (September 15, 2015). "RNA-Based Therapeutics and Vaccines". Genetic Engineering News.
External links
| Wikimedia Commons has media related to mRNA. |
Life of mRNA Flash animation
RNAi Atlas: a database of RNAi libraries and their target analysis results
miRSearch: Tool for finding microRNAs that target mRNA
How mRNA is coded?: YouTube video